







Russian Federation
Russian Federation
UDC 63
CSCSTI 69.01
Russian Classification of Professions by Education 06.06.01
Russian Library and Bibliographic Classification 472
Russian Trade and Bibliographic Classification 64
In the article, an analysis of so-iuy mullet (Planiliza haematocheila) natural reproduction conditions was performed in the Sea of Azov over the past 17 years. The optimal and critical temperatures and salinity of the water were determined for the effective spawning of so-iuy mullet as well as the influence of these factors on its early ontogenesis, the dependence of the migration periods and spawning of producers. An earlier spawning was observed in estuaries and bays.
temperature, salinity, maturation, spawning, ovulation, ontogenesis
Введение
Кефали с древних времен являются ценными объектами морского промысла в Азово-Черноморском бассейне. В настоящее время из обитающих здесь шести видов промысловое значение имеют вселенец пиленгас Planiliza haematocheila = Liza haematocheilus (Temminck & Schlegel, 1845) и три аборигенных черноморских вида: лобан Mugil cephalus (Linneus, 1858), сингиль Liza aurata (Risso, 1810), остронос Liza saliens (Risso, 1810).
В теплое время года аборигенные виды кефалей остронос, сингиль и лобан заходят через Керченский пролив в Азовское море, где нагуливаются, а затем образуют промысловые скопления и мигрируют для нереста или на зимовку в Черном море [2].
По определению Ю.Г. Алеева [1] Черное море является краем ареала черноморских кефалей, где из-за значительного снижения температуры воды в зимний период эти теплолюбивые рыбы, центр ареала которых – субтропические и тропические зоны, не всегда находят оптимальные условия для зимовки, роста и реализации своих высоких воспроизводительных способностей.
Воздействие неблагоприятных внешних условий в отдельные периоды жизненного цикла, определяет численность этих рыб и формирование поколений различной мощности [3; 4]. В конце 50-х годов прошлого столетия формирование в течение ряда лет малоурожайных поколений привели к снижению запасов и депрессивному состоянию популяций черноморских кефалей.
Появление в Азово-Черноморском бассейне дальневосточного вида кефали пиленгаса – результат успешной акклиматизации. В новых условиях проявились высокие показатели его экологической пластичности, темпов роста и способности к успешному естественному воспроизводству в условиях довольно широкого диапазона солености.
В отличие от местных видов, для которых характерны продолжительные зимовальные миграции к берегам Кавказа и Южного Крыма, эта кефаль благополучно зимует в Азовском море, и только во время значительного похолодания образует плотные скопления в распресненных районах моря и в реках (даже в тени небольших мостов).
Первые косяки жизнеспособной молоди пиленгаса в Азовском море отмечались в конце 80-х годов, при их выходе из Молочного лимана [5]. Наиболее многочисленное (высокоурожайное) поколение появилось в 1989 г., молодь наблюдали вдоль всего побережья Азовского моря, а осенью, после похолодания, массовые скопления активно питающихся сеголетков отмечали у берегов Керченского пролива.
Сформировавшиеся в разных районах Азово-Черноморского бассейна самовоспроизводящиеся популяции пиленгаса достигли промысловой численности, и в 1993 г. был разрешен их лов [6]. Численность популяций постепенно увеличивалась, в 2005 г. запас пиленгаса в Азовском море составлял около 62 тыс. т [7]. Однако с 2009 г. его промысловый запас стал постепенно уменьшаться и к 2015 г. составил 2,6 тыс. т (по оценкам специалистов АзНИИРХ) [6].
В настоящий период азовская популяция пиленгаса переживает резкое снижение численности [8]. Вероятно, что для этого вида, также, как и для черноморских кефалей, характерно формирование в отдельные годы урожайных и неурожайных поколений. В связи с этим необходимы дальнейшие исследования особенностей биологии и экологии нереста вселенца, с целью изучения влияния некоторых абиотических условий на эффективность естественного воспроизводства пиленгаса азовской популяции. В ходе исследования решались следующие задачи: 1 – проанализировать динамику температуры в нерестовый период пиленгаса в Азовском море и Керченском проливе в течение последних 17 лет и ее влияние на сроки нерестовых миграций производителей; 2 – исследовать влияние солености Азовского моря на ранний онтогенез и эффективность нереста пиленгаса.
Материал и методы
Материалом послужили результаты ежегодных наблюдений (начиная с 2002 г.) за изменением экологических факторов и физиологического состояния производителей пиленгаса азовской популяции в течение нерестовых миграций в Черное море через Керченский пролив и в Молочный лиман (в 2002, 2003, 2005 гг.). Представлены также результаты экспериментов по созреванию рыб, инкубации икры и развитию предличинок в разных экологических условиях. В экспериментах комбинации солености и температуры соответствовали условиям, наблюдаемым во время нерестового периода в Азовском и Черном морях, а также во время черноморских сгонных явлений. Работы выполнялись на научно-исследовательской базе ЮгНИРО «Заветное», на Молочном лимане (п.г.т Кирилловка) и на КНП ЮгНИРО в Казантипском заливе и в Керченском проливе.
Изучаемая нами популяция пиленгаса зимует в Азовском море. Гонады, в зависимости от температурных условий, осенью и в зимний период находятся на II, II-III или III стадиях зрелости. Весной, при повышении температуры до 6-8˚C, рыба начинает питаться, и развитие половых клеток заметно ускоряется [9]. Самки с гонадами, достигшими IV стадии зрелости, мигрируют к местам нереста. Нерест пиленгаса азовской популяции отмечается как в солоноводных лиманах, заливах и других районах бассейна Азовского моря, так и в черноморских водах.
Сроки миграций и нереста производителей в отдельные годы существенно варьируют и, вероятно, зависят от термики вод Азовского моря и Керченского пролива. Как известно, интенсивность их прогрева обусловлена климатическими условиями в зимний период [10].
Например, весной 2002 г., наступившей после достаточно холодной зимы, ход производителей пиленгаса в Молочный лиман наблюдался уже в начале мая. Температура лиманной воды в канале, соединявшем лиман с морем, колебалась в пределах 16-18ºС, соленость – 20-22‰, в Азовском море – 15-16ºС и 10‰, соответственно. Средний диаметр ооцитов у рыб, заходивших в лиман, изменялся в пределах 487-620 мкм (табл. 1). Число рыб с ооцитами, достигшими дефинитивного состояния (диаметром не менее 600 мкм) в этот период составляло более 35%. Нерестовый сезон 2002 г. характеризовался высокой эффективностью нереста и численностью поколения, о чем свидетельствует положительная динамика вылова в последующие годы [11].
Таблица 1. Размеры ооцитов самок пиленгаса в начале нерестового хода в Молочный лиман в 2002 году
|
Дата |
Температура в Азовском море, ºС |
Диаметр ооцита, мкм |
|
2.05 4.05
5.05 |
17 17 17 17 17 17 17 16,5 |
524,35±11,45 515,54±10,29 487,5±13,8 602,2±11,15 600,74±11,36 505,0±18,65 620,19±3,6 552,5±9,13 |
В 2003 г. после таяния льда в Азовском море благоприятные условия для созревания гонад пиленгаса и нереста в лиманах Северного Приазовья сложились в начале мая. Достаточно быстро прогревались и воды Керченского пролива (рис. 1, а), уже в середине мая здесь появились производители пиленгаса. Однако, как видно на рисунке, сгонные явления, возникшие в Черном море в июне, заметно отражались на динамике температуры в Керченском проливе. Как показали наши наблюдения, при снижении температуры до 12-13ºС нерестовый ход производителей прерывался и возобновлялся только при повышении температуры до 15ºС.
В 2006 г. отмечалась одна из самых суровых и продолжительных зим в Азово-Черноморском бассейне за последние два десятилетия [10], однако весной Азовское море и Керченский пролив прогревались достаточно интенсивно (рис. 1, б). Первые косяки пиленгаса появились в проливе уже 13-15 мая, когда температура в обоих водоемах достигла 12ºС. Сгонные процессы в Черном море, вероятно, оказывали влияние на термический режим не только Керченского пролива, но и там, где температура воды в течение всего июня была ниже 24ºС. Следовательно, благоприятные условия для созревания производителей сохранялись в течение всего нерестового периода пиленгаса.
В течение последнего десятилетия наблюдались лишь две относительно холодные зимы: 2011-2012 и 2012-2013 годы. Соответственно, нерест 2012 и 2013 гг. проходил в сравнительно благоприятных условиях. Производители мигрировали в азовские лиманы и заливы в первой декаде мая. После устойчивого повышения температуры до 24ºС, часть производителей азовской популяции пиленгаса, нерестящаяся в более поздние сроки, по мере созревания, выходила на нерест в Черное море, нерестовый ход продолжался до середины июня.
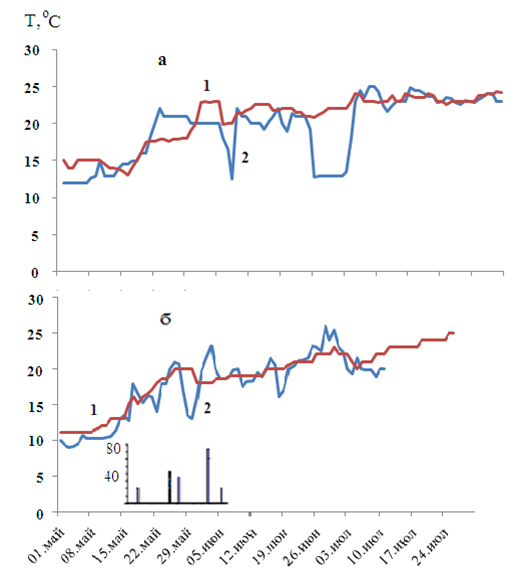
Рисунок 1. Температура в Азовском море (1) и Керченском проливе (2): а – в 2003 году; б – в 2006 г. (на диаграмме: количество самок (%) в улове с признаками резорбции ооцитов)
Мягкие зимы с ранней и затяжной весной отмечались в 2004 и 2005 годах. Нерестовый ход пиленгаса в Молочный лиман наблюдался только с середины мая. В течение июня температура воды в Азовском море в эти годы не превышала 23ºС (рис. 2) и была благоприятной для созревания половых клеток пиленгаса до конца июля.
Как следует из представленных данных, сгонные явления в Черном море в мае и июне также оказывают влияние на тепловой фон Керченского пролива и Азовского моря (рис. 1-3) и, соответственно, на динамику нерестового хода и продолжительность нереста кефалей. Из-за сгонных явлений (апвеллинга), возникающих в Черном море, в Керченском проливе отмечаются резкие перепады температуры. Это отражается на интенсивности нерестового хода производителей. При охлаждении пролива до 12-13ºС нерестовый ход прекращается.
В последующие три года сгонные явления отмечались в июне ежегодно (рис. 3). При этом в мае Керченский пролив прогревался достаточно интенсивно, благодаря чему пиленгас появлялся здесь уже в конце апреля и начале мая. Так в 2008 г. после более суровой зимы первые уловы пиленгаса в проливе отмечались 28 апреля, при этом температура была выше, чем в Азовском море: максимальная температура 16,5ºС отмечена 10 мая, в Азове в этот день она не превышала 12ºС.
Как показали исследования, при резком снижении температуры воды с 18-22°C до 12-13°C, даже на короткое время – на 2-3 суток, у всех опытных рыб наблюдалась резорбция желтковых ооцитов. Обращает на себя внимание то, что в первую очередь дегенеративным изменениям подвергаются самые крупные в гонадах желтковые ооциты, близкие к дефинитивным размерам.
При резком снижении температуры воды с 18-22°С до 12-13°С, даже на короткое время – на 2-3 суток, у всех опытных рыб наблюдалась резорбция желтковых ооцитов. Обращает на себя внимание то, что, в первую очередь, дегенеративным изменениям подвергаются самые крупные в гонадах желтковые ооциты, близкие к дефинитивным размерам. При недолгом действии неблагоприятного фактора уцелевшие желтковые клетки, как правило, созревают нормально. Однако зрелая икра, полученная от сохранившихся ооцитов, характеризуется меньшей сухой массой, чем полученная от одноразмерных ооцитов без признаков резорбции. Это может отражаться на рыбоводно-биологических показателях ранних личинок и их жизнеспособности [12].
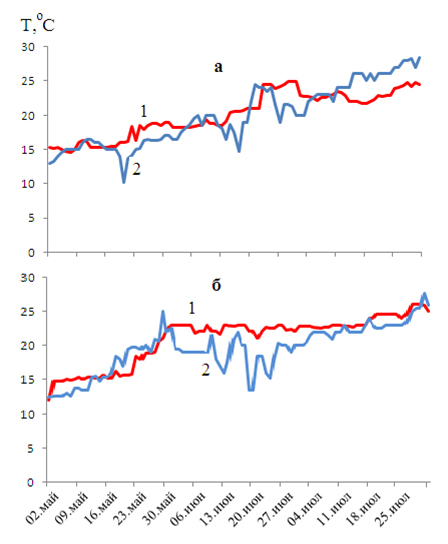
Рисунок 2. Температура в Азовском море (1) и Керченском проливе (2) в 2004 (а) и 2005 (б) годах
Дегенеративные изменения в созревающих ооцитах кефалей и нарушения на завершающих фазах их созревания, наступают также при повышении температуры до 24ºС и выше. В 2005 г. такая ситуация наблюдалась в Молочном лимане после 20 июня. В гонадах у рыб, выходящих из лимана в этот период, отмечались остатки зрелых ооцитов с разной степенью резорбции, что свидетельствовало о прошедшем нересте. Кроме того, в яичнике этих же рыб находилось еще значительное количество желтковых ооцитов, близких к дефинитивному состоянию, часть которых имела признаки дегенерации. Вероятно, неблагоприятные температурные условия не способствовали созреванию большей части ооцитов – генерации клеток, предназначенных для вымета в текущем сезоне. Однако у таких рыб, при попадании в нерестовые условия, возможен повторный нерест. В наших экспериментах после гормонального инъецирования созревания рыб, находившихся в подобном состоянии, полученные овулировавшая икра и личинки характеризовались хорошим рыбоводным качеством.
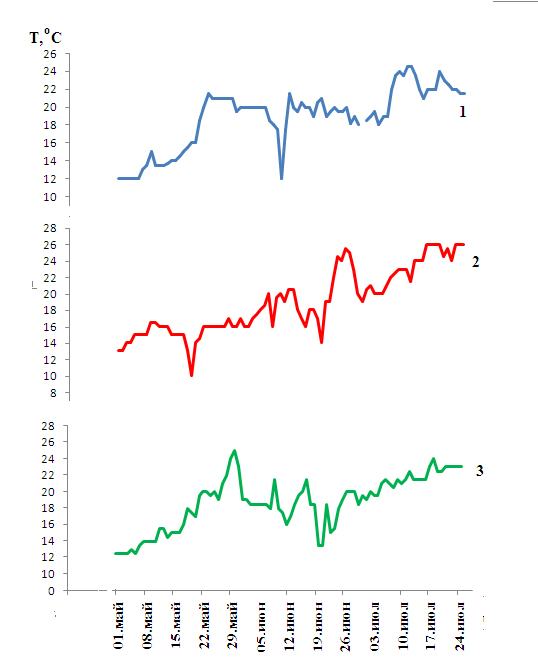
Рисунок 3. Динамика температуры в Керченском проливе в 2007 (1), 2008 (2), 2009 (3)
После теплых зим 2014-2018 гг. также отмечались холодные весенние месяцы и отсутствие массового нерестового хода пиленгаса в Керченском проливе. В Азовском же море в эти годы нерестовые условия наблюдались в течение всего июня (табл. 2, 3, рис. 4). Лишь в июне 2019 г., несмотря на теплую зиму, температура воды в прибрежной зоне практически во всех районах Азовского моря стала интенсивно повышаться, и уже 7 июня достигла 24ºС (см. табл. 3), что и обусловило начало мощного нерестового хода рыбы в Керченском проливе.
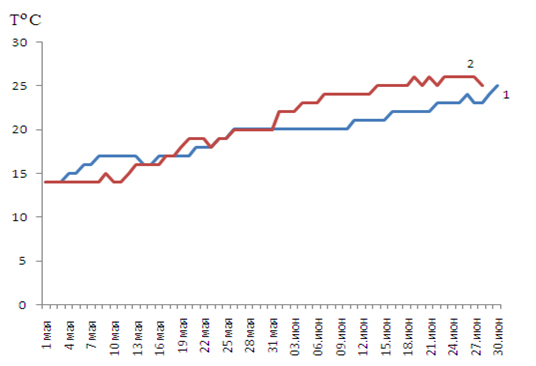
Рисунок 4. Температура в Азовском море: 1 – в 2018, 2 – в 2019 годах
Таким образом, начало нерестового хода пиленгаса через Керченский пролив и его интенсивность зависят от степени прогревания Азовского моря и пролива. Сигнальное значение для начала миграций, по-видимому, имеет повышение температуры поверхностных слоев этих водоемов с 12 до 15ºС (табл. 2) и вынос из пролива в Азовское море водных масс более высокой солености.
Таблица 2. Даты повышения температуры до 15 ºС в прибрежных районах Азовского моря и Керченского пролива
|
Годы |
Районы исследований |
Характеристика зимнего периода |
||
|
пгт Кирилловка |
с. Мысовое |
Керченский пролив (первые уловы пиленгаса) |
||
|
2009 2010 2011 2012 2013 2014 2015 2016 2017 2018 2019 |
18.05 20.05 22.05 5.05 3.05 11.05 14.05 10.05 19.05 5.05 11.05 |
18.05 20.05 22.05 5.05 3.05 13.05 14.05 10.05 26.05 6.05 12.05 |
18.05 20.05 22.05 5.05 3.05 13.05 14.05 10.05 24.05 10.05 16.05 |
мягкая зима умеренная умеренная холодная холодная умеренная мягкая мягкая мягкая мягкая мягкая |
Как следует из представленных данных, продолжительность и начало нерестовых миграций пиленгаса существенно варьируют в разные годы, и зависят от интенсивности прогревания Азовского моря и Керченского пролива.
В соленые лиманы Северного Приазовья пиленгас также начинает заходить после их прогревания до 15ºС. Завершающие процессы созревания ооцитов и овуляция зрелых яиц происходит в довольно узком интервале температур – 16-23ºС.
Результатов наших исследований показали, что наиболее благоприятные условия для нереста пиленгаса в Азово-Черноморском бассейне складываются после суровых и умеренных зим: Азовское море и Керченский пролив прогреваются более интенсивно и синхронно, миграции производителей к местам нереста начинаются в конце апреля-начале мая, нерест раннесозревающей части популяции азовского пиленгаса проходит дружно в интенсивно прогревающихся азовских лиманах. После повышении температуры до 24ºС нерестовая активность у рыб снижается; производители, созревающие в более поздние сроки, и имеющие гонады IV, III-IV стадий зрелости мигрируют через Керченский пролив в Черное море, в более прохладные и соленые воды.
В то же время, после мягких зим, как известно, часто наступает ранняя, холодная и затяжная весна [10]. В мае и июне температура Азовского моря повышается медленно и часто вплоть до конца июня-начала июля не достигает 24ºС (табл. 3). Мелководные лиманы прогреваются до 24ºС достаточно быстро, что приводит к нарушениям созревания ооцитов и развития эмбрионов и ранних личинок. Соответственно, эффективность нереста снижается.
Таблица 3. Даты устойчивого повышения температуры до 24ºС в Азовском море
|
Годы |
Районы исследования |
||
|
Мысовое |
Кирилловка |
Таганрогский залив |
|
|
2009 2010 2011 2012 2013 2014 2015 2016 2017 2018 2019 |
25.06 13.06 17.06 13.06 19.06 5.07 16.06 21.06 28.06 26.06 7.06 |
25.06 13.06 17.06 12.06 19.06 4.07 16.06 19.06 28.06 22.06 7.06 |
11.06 12.06 19.06 12.06 18.06 12.07 11.06 20.06 28.06 17.06 30 05 |
Нормальное завершение созревания ооцитов и овуляция качественной зрелой икры происходят в диапазоне 15-23ºС. Как показано ранее, при температуре выше 24ºС созревание самок часто заканчивается тотальной резорбцией половых клеток, а предличинки, на этапах вылупления и перехода на внешнее питание, оказываются ослабленными из-за интенсивного расходования питательных веществ яйца [12]. В теплой азовской воде низкой солености плавниковая кайма предличинок часто поражается бактериями.
В случае сгонных явлений и возникновения апвеллинга нерест может происходить в Керченском проливе. В случае его значительного охлаждения рыба вынуждена оставаться в Азовском море и искать для нереста наиболее благоприятные места, адаптируясь к новым условиям.
В связи с осолонением Азовского моря до 13-14‰, наблюдаемом в последние годы, нерест пиленгаса отмечается даже в самых опресненных некогда районах: Таганрогском, Бейсугском заливах и др. По данным В.П. Надолинского [13], икра пиленгаса, нерестящегося в разных районах Азовского моря, характеризуется значительной вариабельностью диаметров икринки (0,6-1,0 мм) и жировой капли (0,4-0,65 мм). Относительный объем жировой капли, рассчитанный нами по средним показателям, приведенным автором, составляет 12,95-18,82%.
Полученные данные свидетельствуют о тенденции увеличения жировой капли и ее относительного объема у рыб, нерестящихся в Азовском море. Как известно, в раннем онтогенезе многих видов рыб жировая капля выполняет в основном гидростатическую роль и определяет нейтральную плавучесть икры. Увеличение относительного объема жировой капли до 19% обеспечивает икре пиленгаса возможность сохранять положительную плавучесть в азовской воде. Однако это свидетельствует о снижении диаметра икры, а, следовательно, и запасов энергетических и пластических веществ в яйце, что может отразиться на жизнеспособности личинок.
Таким образом, в связи с мелководностью азовских лиманов и заливов происходит достаточно быстрое их прогревание. Уже в начале лета отмечаются критические показатели температуры, вызывающие резорбцию ооцитов и нарушение завершающих процессов созревания яйцеклеток у пиленгаса. В результате, нормальный нерест в лиманах Азовского моря может наблюдаться лишь у небольшого количества производителей пиленгаса, созревающих в более ранние сроки, а в самом Азовском море – только у рыб, продуцирующих мелкую икру, способную сохранять положительную плавучесть в азовской воде. Позднесозревающие рыбы, а также их потомство оказываются в неблагоприятных условиях из-за повышающейся температуры. Личинки характеризуются низкой жизнеспособностью, в связи с повышенными энергетическими тратами во время эмбрионального и раннего личиночного развития (на стадии предличинки), и вероятно, бактериальной обсемененностью в теплой азовской воде, что приводит к нарушениям процессов перехода на внешнее питание, заполнения плавательного пузыря, пищеварения. Кроме того, в теплой воде быстро расходуются запасы желточного мешка и жировой капли, такая ослабленная малоподвижная личинка характеризуется, как правило, низкой жизнеспособностью.
Выводы
1. После мягких зим, как правило, наступает затяжная холодная весна, способствующая медленному прогреванию Азовского моря в мае и июне (нерестовый период пиленгаса). Это приводит к более позднему нересту пиленгаса в лиманах Азовского моря, растянутости нерестового периода и снижению интенсивности его нерестового хода через Керченский пролив.
2. В лиманах и заливах Азовского моря при температуре 16-23ºС и солености 17-23‰ нерестятся раннесозревающие производители пиленгаса. При повышении температуры воды на местах нереста до 24ºС и выше наблюдаются нарушения созревания, характеризующиеся резорбцией половых клеток производителей, частичной овуляцией или выметом перезревших яйцеклеток. Эффективный нерест пиленгаса непосредственно в Азовском море может происходить при температуре не выше 24ºС. У рыб, созревающих при более высокой температуре, также отмечаются нарушения оогенеза и процесса овуляции половых клеток.
3. Сгонные явления в Черном море оказывают существенное влияние на поведение производителей пиленгаса: при снижении температуры воды в Керченском проливе до 9-12ºС нерестовый ход прекращается, у рыб, попавших в зону низких температур, из-за резкого изменения температуры отмечается резорбция половых клеток, а эмбрионы и личинки впадают в анабиоз.
4. Повышение солености Азовского моря в последние годы способствует созданию благоприятных условий для нереста части производителей пиленгаса в некоторых открытых его районах. Однако зрелая икра этих рыб характеризуется более мелкими размерами, и при повышении температуры у эмбрионов и личинок происходит ускоренное расходование питательных веществ, что влияет на их жизнеспособность и устойчивость к бактериальной флоре и условиям среды.
1. Aleev Yu.G. O nekotoryh zakonomernostyah rosta ryb / Yu.G. Aleev // Voprosy ihtiologii.- 1966. - vyp. 6. - S. 75-95.
2. Diripasko O.A. Ryby Azovskogo morya / O.A. Diripasko, L.V. Izergin, K.V. Dem'yanenko // Izd-vo OOO «NPK» Inter - MN, 2011. - 288 s.
3. Timoshek N.G. Harakteristika sostoyaniya zapasa chernomorskih kefaley v 1961 godu, utochnenie vozmozhnogo ih vylova v 1962 godu i predvaritel'nyy prognoz na 1963 god / N.G. Timoshek // Trudy AzCherNIRO. - 1961. - 25 s.
4. Timoshek N.G. Kefali / N.G. Timoshek, R.M. Pavlovskaya // Syr'evye resursy Chernogo morya. - M.: Pisch. prom-t', 1979. - S 175-208.
5. Yanovskiy E.G.Formirovanie promyslovoy populyacii pilengasa / E.G. Yanovskiy, L.V. Izergin // Rybnoe hozyaystvo, 1995. - № 4. - S. 42-43
6. Balykin P.A. Rezul'taty vseleniya pilengasa v Azovo-Chernomorskiy basseyn / P.A. Balykin, A.V. Starcev // Prirodnye resursy, ih sovremennoe sostoyanie, ohrana, promyslovoe i tehnicheskoe ispol'zovanie: materialy VIII Vserossiyskoy nauchno-prakticheskoy konferencii, posvyaschennoy 75-letiyu rybohozyaystvennogo obrazovaniya na Kamchatke (12-14 aprelya 2017 g.): v 2 ch. / otv. za vyp. N.G. Klochkova. - Ch. I. - Petropavlovsk-Kamchatskiy: KamchatGTU, 2017. - 34-37.
7. Besedin V.B., Rekov Yu.I. Sovremennyy etap stanovleniya populyacii pilengasa v Azov-skom more / V.B. Besedin, Yu.I. Rekov // Sb. nauch. tr. AzNIIRH «Osnov. probl. ryb. hoz-va i ohrany rybohoz. vodoemov Azovo-Chernomorskogo basseyna». - Rostov n/D., 2003. - S. 181-188.
8. Kozhurin E.A. Pilengas: akklimatizaciya, biologicheskiy vzryv, depressiya i perspektivy promysla / E.A. Kozhurin - Rybnoe hozyaystvo, 2018. - №1. - S. 92-94.
9. Fiziologicheskoe sostoyanie proizvoditeley pilengasa v period migracii cherez Kerchenskiy proliv / N.I. Kulikova, A.F. Bulli, L.G. Gnatchenko, I.I. Pisarevskaya, V.N. Fedulina, L.I. Bulli // Trudy YugNIRO. - 1996. - t.42 - S. 210-216
10. Lavrova O.Yu. Ledovaya obstanovka v Kerchenskom prolive v tekuschem stoletii. Retrospektivnyy analiz na osnove sputnikovyh dannyh / O.Yu. Lavrova, M.I. Mityagina, A.G. Kostyanoy // Sovremennye problemy distancionnogo zondirovaniya Zemli iz kosmosa. 2017. - T. 14. № 2. - S. 148-166.
11. Kozhurin E.A. Dinamika ulovov promyslovyh ryb Kryma v Chernom more / E.A. Kozhurin, V.A. Shlyahov, E.P. Gubanov // Tudy VNIRO, T. 171. - 2018. - S.157-169.
12. Bulli L.I. Osobennosti sozrevaniya i harakteristika ikry pilengasa, migriruyuschego cherez Kerchenskiy proliv / L.I. Bulli // Rybnoe hozyaystvo Ukrainy - 2004. - № 7 (spec. vypusk po mater. II Mezhdun. n-prakt. konf. «Morskie tehnologii: problemy iresheniya - 2004») - S. 92-97.
13. Nadolinskiy V.P. Dinamika raspredeleniya morskih nerestilisch i adaptaciya pilengasa k usloviyam razmnozheniya v Azovskom more / V.P. Nadolinskiy // Voprosy rybolovstva - 2008. - T. 9. №4 (36). - S. 807-814.